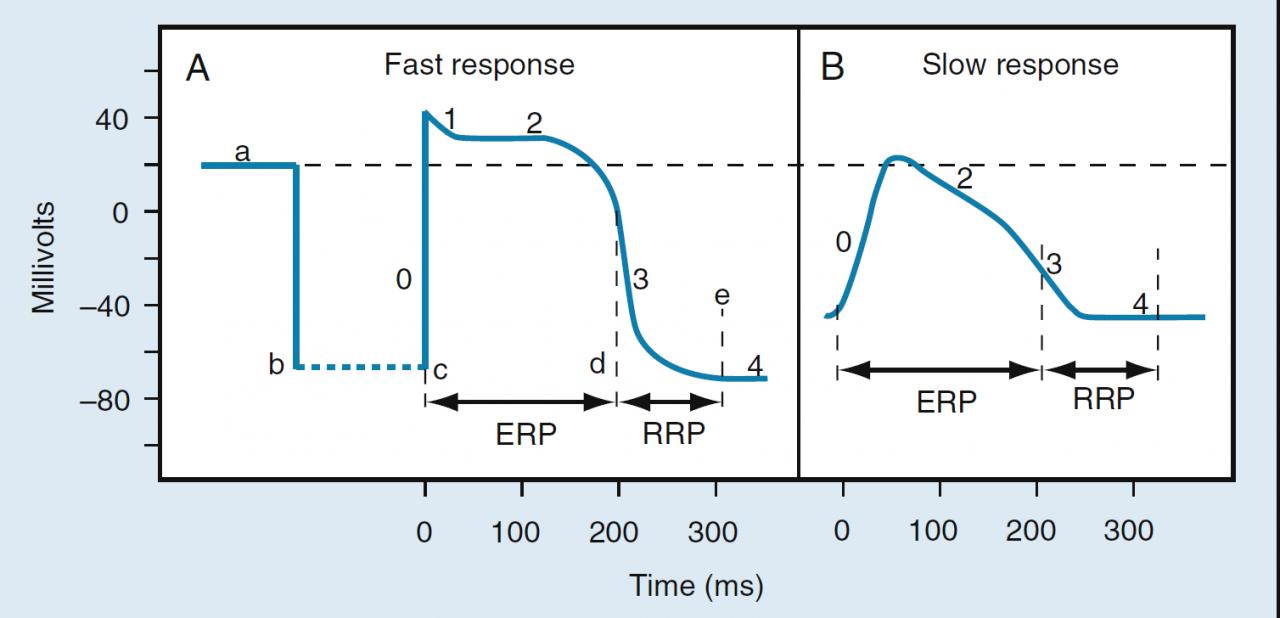
Bonsoir tout le monde, j'ai une question qui se rapporte au tissu nodal, nous avons dit que les cellules nodales sont restées à l'état embryonnaire et de ce fait les disques intercalaires ne sont pas encore différenciés alors comment ça se fait que les cellules du noeud auriculo ventriculaire ont la vitesse de conduction la plus faible mais ceux de purkinje ont celle la plus rapide, vu que toutes les deux appartiennent à ce tissu nodal !
Merci pour vos réponses.
-----