
Comment est régénéré le succinate ?
(Complément indispensable pour une réponse complète !)
(Et "pyruvate + CoA-SH -> Acetyl CoA avec libération de CO2" ne paraît pas équilibré, il y a un H manquant, non?)
-----

Comment est régénéré le succinate ?
(Complément indispensable pour une réponse complète !)
(Et "pyruvate + CoA-SH -> Acetyl CoA avec libération de CO2" ne paraît pas équilibré, il y a un H manquant, non?)
Dernière modification par Amanuensis ; 04/04/2013 à 11h12.
Pour toute question, il y a une réponse simple, évidente, et fausse.
C'est fort possible qu'il y ait un H manquant, c'est courant en biochimie de ne pas tous les mettre. Il y en a tellement de disponible dans la cellule de toute façon.
Sinon c'est la réaction très classique SuccinylCoA -> succinate couplée à de l'ADP + Pi qui fournit l'ATP. Je te renvoie au cycle de Krebs, c'est la même réaction.
Sûr. Sauf qu'ici c'est justement le décompte des H qui m'intéresse.Envoyé par Alhec
Oui, et cela implique une molécule d'eau, dont l'oxygène régénère celui cédé là:Sinon c'est la réaction très classique SuccinylCoA -> succinate couplée à de l'ADP + Pi qui fournit l'ATP. Je te renvoie au cycle de Krebs, c'est la même réaction.
Dans mon message #30, je n'étais pas entré dans les détails du passage par le Succinyl-CoA, mais avais indiqué la molécule d'eau et les H.L'oxygène en plus vient du succinate.
En détails :
pyruvate + CoA-SH -> Acetyl CoA + CO2 + H
Acetyl CoA + succinate -> Succinyl CoA + acétate
Succinyl CoA + H2O -> succinate + CoA-H + H
D'où le bilan que j'avais indiqué.
À ce stade, on a "en trop" NADH + H+ venant de l'étape 1/2 glucose -> pyruvate, et 2 H venant de l'étape suivante. J'ai fais l'hypothèse que le H2 généré par l'hydrogénosome étaient les H de la seconde étape. Non ?
----
Désolé d'être têtu, je cherche juste à trouver un bilan comptable propre, avec les centimes. Je ne prétends pas que celui que j'indique un soit le bon, mais j'en cherche un complet...
Dernière modification par Amanuensis ; 04/04/2013 à 12h24.
Pour toute question, il y a une réponse simple, évidente, et fausse.
Honnêtement, je n'en sais rien et je ne sais pas si quelqu'un s'est déjà penché sur la question.
Par contre, qu'est-ce qui t'intéresse tant dans le décompte des H? Dans une chaîne respiratoire je comprendrais, mais ici?
Mon interrogation est présente dès le début: est-ce que l'utilisation du glucose pour fournir 2 ATP est optimale ? Il y a deux NAD+ -> NADH + H+ qui sont "jetés" juste pour réduire le pyruvate et rejeter de l'éthanol ou de l'acide acétique, et la seule justification que j'ai trouvée est le recyclage des NAD+.
Comme j'ai cru voir une mention de "0 à 2 ATP" supplémentaire quelque part, je cherche ce qui pourrait faire mieux que ces 2 ATP de base. Et la question semble bien être liée au recyclage de NADH + H+ en NAD+, c'est à dire quoi faire de ces "2 H" en trop...
Dernière modification par Amanuensis ; 04/04/2013 à 13h22.
Pour toute question, il y a une réponse simple, évidente, et fausse.
Le TPP ou thiamine pyrophosphate est un coenzyme qui est réduit lors de la décarboxylation oxydative du pyruvate en acétate (acétyl-phosphate pour être exact). L'énergie de cette oxydation est transférée en partie dans une liaison ester phosphorique avec l'acétate formé, tandis que les protons (et électrons) sont envoyés sur le TPP (qui devient donc TPPH2). L'hydrogénase est l'enzyme qui va ensuite réoxyder le TPPH2 en TPP, en libérant le dihydrogène qui s'échappe dans le milieu extérieur au même titre que le CO2 formé à la première étape. Ces deux gaz ayant un excellent pouvoir de diffusion, ils se "diluent" très vite, déplaçant l'équilibre de la réaction ce qui la favorise d'autant plus.
Auriez-vous quelques liens vers cette respiration soufrée ?
OK. Quand on parle de respiration, il s'agit de ré-oxyder les co-enzymes. Si elle est qualifiée d'anaérobie, c'est parce qu'on utilise une autre molécule que le O2. Implication d'enzymes différentes, et bilan ATP différent. J'ai un peu oublié le détail de la respiration nitrate et nitrite, je vais donc revoir ça en détail avec les sulfates et sulfites
J'avais la suspicion qu'une respiration carbonate ou CO2 puisse exister mais j'ignorais que c'était la voie des méthanogènes !Merci

Question : l'hydrogénase de Clostridium est-elle libre dans le cytoplasme ou se situe-t-elle dans la membrane plasmique à l'image de l'oxydase chez Pseudomonas ?
Pas avec détails. Le cas O2 est décrit partout, les autres c'est plus difficile.
À ce que je crois comprendre, c'est la même chaîne, la seule différence est l'oxydant pour NADH + H+ -> NAD+ (par exemple), qui se couple avec SO4= -> SO3 + H2O (j'imagine), ou S -> SH2.
J'aimerais trouvé les bilans détaillés, la différence doit être le nombre de H+ qui passent de l'autre côté de la membrane.
Pour toute question, il y a une réponse simple, évidente, et fausse.
J'ai trouvé cette page avec pas mal de choses mais pas la respiration soufrée ^^;
Peut-être y trouvera-t-on quelques équations de synthèse ATP supplémentaires ?
On a oublié la synthèse ATP par la pompe à protons je crois.
Tant qu'on y est, comment fonctionne les hydrogénases? Ne devraient-elles pas consommer de l'ATP? Il me semble que les électrons venant du NADH ne peuvent pas réduire H+, donc il faut "forcer" la réaction? Ou bien je me trompe (les redox, ça a jamais été mon fort).
Concernant la formique hydrogène lyase, il ne semble pas y avoir de consommation ATP.
Concernant les hydrogénases, on a réversibilité de réaction :
2H+ + 2e- <==> H2
Aucune consommation ou production d'ATP par ces enzymes de mentionner -- sans doute à imputer à la réversibilité réactionnelle.
(annulé, erreur)
Pour toute question, il y a une réponse simple, évidente, et fausse.
Je ne comprend pas trop la question de départ. Si on s'interdit d'utiliser un oxydant, on ne peut tirer aucune énergie d'une molécule de glucose ! (ou alors je n'y connais vraiment rien). Donc la réponse c'est : ZERO molécules d'ATP !
Je confirme que vous ne connaissez rien au métabolisme glucidique.
Non seulement lors de la glycolyse il y a création d'ATP (bilan net : 2 ATP + deux pyruvates), mais en plus le recyclage des co-enzymes "respiratoires", que ce soit par respiration ou par fermentation, va produire là encore des ATP (cf pompe à proton).
PS : Il faudrait par ailleurs définir ce que vous entendriez par "un oxydant", terme bien vague en biochimie.
Je vous rassure, je connais un peu le métabolisme, mais j'avais lu rapidement qu'on n 'avait "pas le droit" d'utiliser un oxydant (hors le NAD+ en est un) ce qui n'avait pas de sens. Mais en fait, il est bien précisé qu'il ne faut pas d'oxydant "extérieur", donc mea culpa je me suis planté.
Mea culpa aussi pour la mauvaise interprétation de votre propos. Dans l'absolu on effectue bien des oxydations (dont celles avec des co-enzymes qui sont ensuite recyclés).
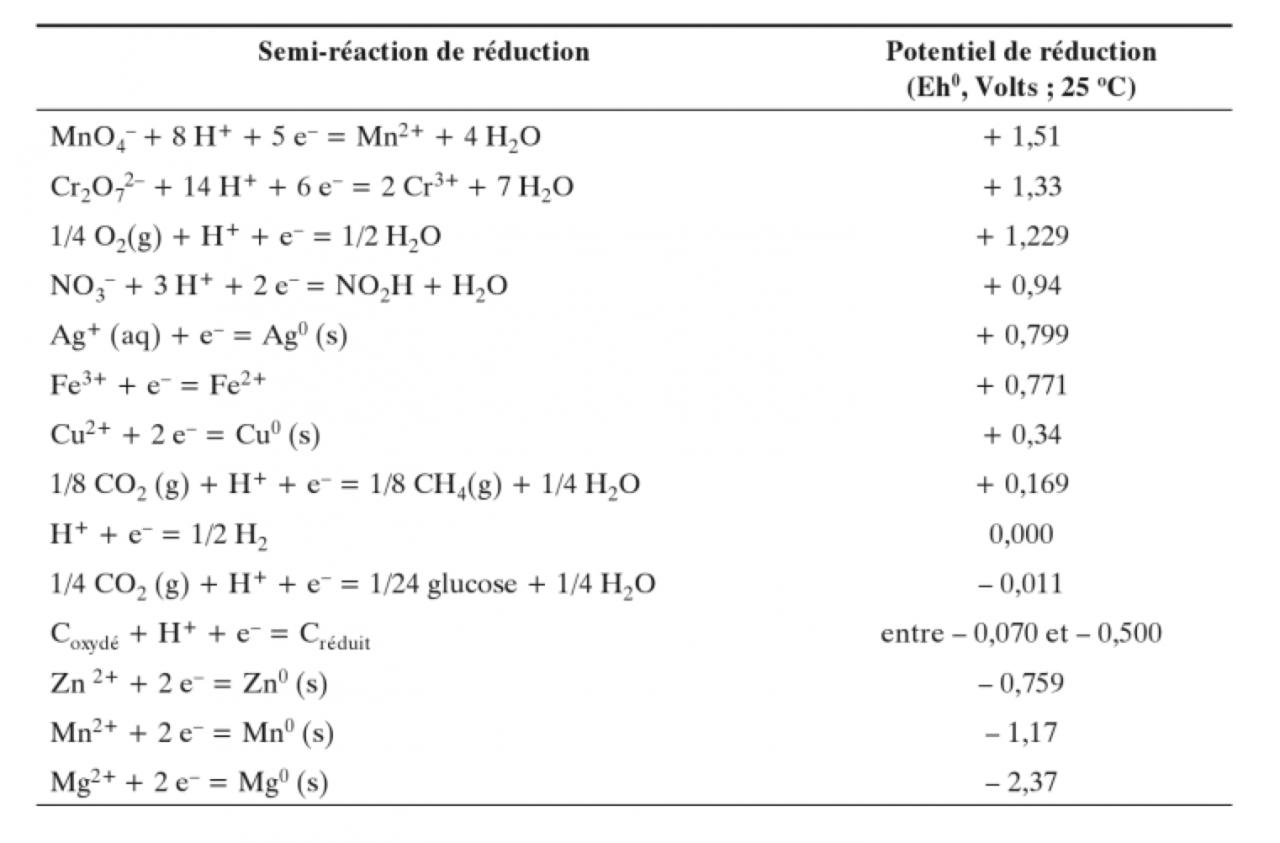
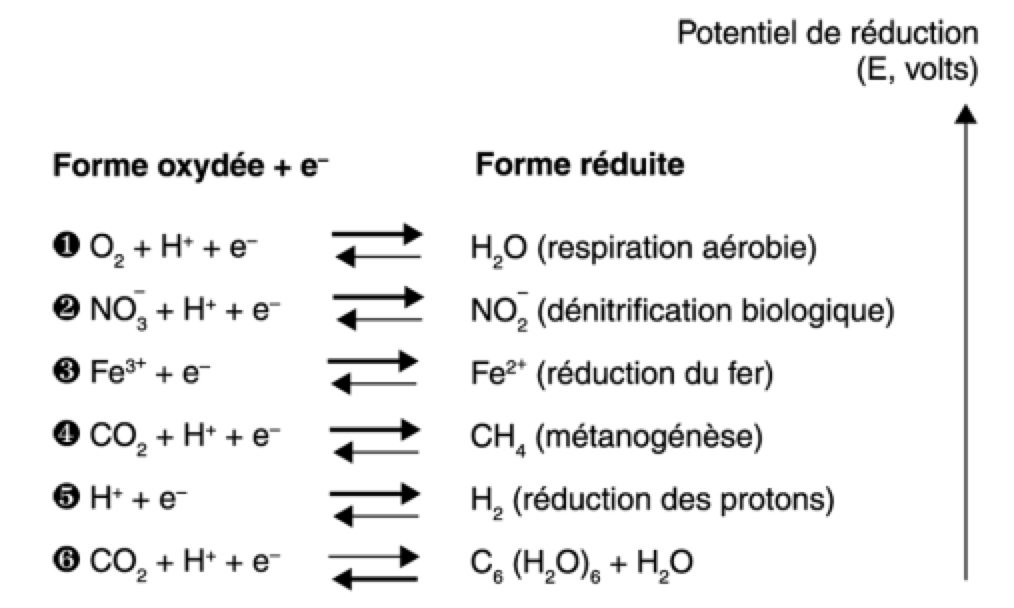
Pour reprendre de manière constructive, voici un tableau résumant les potentiel redox tirés d'un bouquin de microbio (dont les extraits sont disponibles librement sur google books) :
Redox1.jpg
Redox2.png
Ça permet d'avoir une bonne vue d'ensemble sur ce qui est possible ou pas de tirer comme énergie du glucose. Le reste du bouquin a l'air pas mal non plus : Principes de chimie redox en écologie microbienne

{kind=link}
{kind=link}